A.00 Mechanisms: From Simple Beginnings to Hyper-Complexity
A.01 Simple Beginnings |
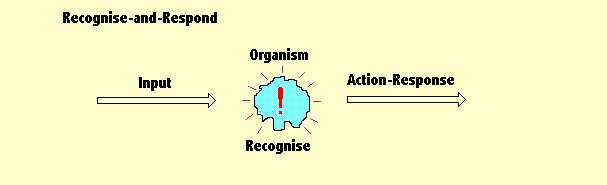
| This diagram shows the behaviour of the simplest animate organism I can think of. A bit of Pond life. A microscopic blob of jelly. |
|
So - when the organism has recognised something (whatever it is), it then does the second thing it can do. It triggers a fixed action-response. That is, it sends a signal to a part of the brain which controls physical actions. The term physical actions covers everything from overt muscular actions, like limb movements, to internal covert actions, like re-arranging data stores in its brain. The things it can recognise and the actions it can take in response, are paired. If it encounters THIS it will respond by triggering THAT. THIS will trigger THAT - always - unless the mechanism undergoes a mutation. Mutations are spontaneous changes to the organism's genes. Genes control chemical reactions inside an organism and mutations are random. Being random those mutations are also erratic. The average rate of mutational change is more or less constant, but there are occasional splurges and times when they do not happen often. The things which can cause mutations are some chemicals, some viruses and nuclear radiation. That is why accidents to nuclear power stations are usually followed for a few years by an increase in the identification of cancers in the local population. In the days before scientists learned how to modify genes deliberately, agriculturists, who wanted the create new types of plants, planted some crops in circles round a source of radiation (like a plug of radio-active Cobalt-60) ... and then they waited, for months. After a while the growing crops were examined. The ones that were suffering some kind of radiation damage - and that means most of them - were discarded. But among those were a small number which had beneficial changes to their genes were used as a seed crop for the next crop - to test for an ability to grow well during a drought, an ability to fight off infections, and other good characteristics. It was a wasteful and inefficient way of producing crops with these desirable characteristics, but it was also the only way they had available before they learned how to do what is called "gene editing". Anyway, at first the range of things an organism could recognise and the range of actions it could trigger were both limited. Evolution, however, over an enormous length of time, gradually extended the range of circumstances that it could recognise and the range of responses it could trigger. This extension was caused by those random mutations. The consequences of behaving in that random way are almost always very bad and sometimes fatal. But sometimes, very very rarely, the results are beneficial. An organism which has these good mutations will survive and reproduce more organisms like itself. And that, it seems, is why a population of organisms will gradually get better and better at surviving in its environment .... provided that environment stays more or less unaltered. And that is the mechanism which operated and ensured that lifeforms survived. And survive, it seems, is what they did and continued to do for three billion years ... changing very slowly. Slowly adding to their capabilites. We call it "natural selection" but in reality there really is no "selection" going on at all. Not, at any rate a form of deliberate selection. Organisms with characteristic behaviour which helps them to survive - do survive (mostly). Everything else does not survive (mostly). Because the mechanism is a bit hit and miss, it takes a very long time indeed to get anywhere. |
|
The diagram does not make any commitment to a particular type of perception. It is a schematic illustration of the jelly blob in action. It provides the basis for the description which follows. |
|
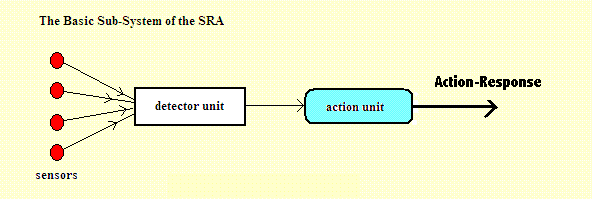
At an early stage of the evolutionary path, these round blobs (on the left) might simply be able to react chemically with some other object. So, in effect, our jelly blob is able only to taste the things it bumps into. Or maybe it doesn't have to really make physical contact. The chemicals it can taste may be dissolved in the water around it. At a slightly later stage those round blobs might correspond to light-sensitive spots which simply tell the difference between daylight and darkness. Too bright? - Go deeper into the water (into darkness). Too dark? - Rise upwards (into the light). Just right? - Don't move. The principle is easily stated. First, examine the group of signals being received (from the left), by whichever type of perceptions are involved. That is the job done by the "Detector Unit". If that unit confirms that the prescribed characteristics are present within that input (within some defined approximation), then a predefined action-response is triggered. That is the job done by the "Action-Unit". It could be a very simple action, or it could at a latter stage be something quite complicated - a sequence of muscle contractions for example which are commands which choreograph the behaviour. But note - there is no choice involved. There is no intellectual calculation. Neither is there any form of memory. All these things will evolve later. Anyway, if the SRA behaves in that simple way, it may then also find itself surrounded by more organisms which are behaving in a similar way. That could be good thing or a bad thing. Our organism might, for example, be able to eat those other organisms. But then again, those other organisms might be able to eat our jelly blob organism. Ah well ... there are probably plenty more like it to keep this story going. After a few aeons of evolution, those light-sensitive spots may have evolved into eyes - eyes with a lens, or with several lenses as in the case of a compound insect eye. So now the blobs may correspond to cells inside an eye (or multiple compound eyes). These cells may be set in a particular pattern. That is, the way they are laid out corresponds (presumably) to the source of the incoming signals. So incoming light is focused on to these retinal cells and the pattern of those signals will be processed. If there is a match between the incoming pattern of signals and the pattern of light-sensitive cells, an output signal is then sent to a detector unit. That is an automatic response. There is no intellectual involvement. Mutations continue. More, and different kinds of perception develop. More acute vision. Sound sensors (sensitive to air vibration). Touch sensors (sensitive to physical contact). Taste receptors (sensitive to chemical reactions with other molecules). Scent receptors (likewise, to airborne molecules). In some cases, especially in insects, the blobs will be structured to correspond to gradients of scented mlecules. Detection of a gradient of molecules will trigger a direction in which to fly. A creature thus endowed, will fly up the gradient to reach a source of that scent, even a very weak gradient. It could be a source of food. Or a gradient of pheromones which could indicate a source of sex. And these complex perceptions are associated with more and more complicated types of action-response. Movement of limbs. Patterns of complicated movement. Emission of sound (sending a warning to others, getting help from parents). Running away (escape from a predator). Our knowledge of how animals tended to behave at these times is limited by the fact that behavioural characteristics do not (normally) leave behind any fossil evidence except for the marks they leave behind in soft mud and similar surfaces. Quote: "The earliest trackways date to over 585 mya [million years ago] and were made by a small worm less than 1cm long, with simple actions. The only "rules" for these actions were: follow the food gradient, do not dig deeply or vertically, and plow straight ahead or in shallow waves with no exploratory side trips. Fascinatingly this is "fossilized behaviour," of a very primitive sort." [Feinberg and Mallatt 2016 p57 (quoting evidence from a 2012 report by Pecoits et al)]. A.02 More Sub-Systems: The Simple Evolution of the SRA |
|
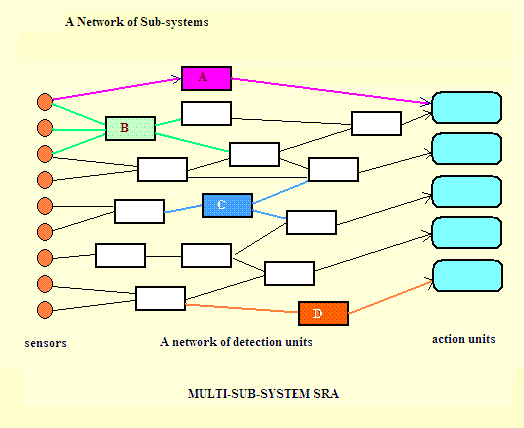
The diagram above illustrates the nature of the SRA, after a relatively short period of evolutionary development. As shown in the diagram, it is a network of sub-systems. Each sub-system is like the one shown in the previous diagram. In this form the SRA has a range of conditions which it can recognise and a range of actions any one of which it can call upon to act as the proposed action-response. |
|
So for all these patterns of behaviour ... the recognition of various conditions which then each trigger an action-response, can be happening simultaneously. They may be very simple, but they have this considerable advantage ... by being effectively simultaneous, (if they are not mutually exclusive) they respond at great speed. After a more prolonged period of evolution the SRA will become a network of a great many thousands or even multiple millions of sub-systems. Each of these subsystems receives input signals from one or more sensors (probably a lot more than one). Those signals are generated by the mechanisms of perception. Some of these are external sensors (eyes, nose, finger-tip sensors, ears and taste buds). But some are internal sensors. These can detect internal conditions like the concentration of glucose dissolved in the blood. We associate a low level of glucose with a condition which we often call "hunger", recognition of which would normally trigger the action-response of eating (if food is available). Now the organisms exhibit a complex range of behaviours. They have a pattern of behaviour which we could call "an escape routine". They can indulge in sexual reproduction. They can prey on smaller organisms. An important new feature will be a memory. A.03 Speed vs Complexity And now our simple but evolving organism runs into another problem. How can it evolve further, to become a very complicated organism indeed, without making itself very much slower? To see my answer to that, see later in this text. For the time being, however, I shall continue to offer suggestions which make it more complex and more sophisticated, but I will defer the explanation of how these complexities could be reorganised. A.04 Secondary Goals and Chain Logic We can expect that primitive organisms would try to eat almost continuously whenever the presence of food is identified. At a later stage, when food is encountered at irregular intervals, we can expect to observe a more complicated pattern of behaviour which takes other considerations into the decision-making process. The current state of the organism's nutrition, is one of those conditions which are taken into account. The rule could be expressed this way ... "If predators are about, and you are not not hungry, then stay hidden". The introduction of that conditional "if" clause, if it is a "good" rule, will increase the complexity of the mechanism, and increases the chances of survival. However, for such an ability to operate effectively the organism must be able to remember how recently it perceived a predator and must be able to recall its current state of nutrition. Initially GOAL conditions will be acquired very slowly, by fortuitous mutations producing characterisics which promote survival. But at those later stages a faster method of acquiring survival-linked characteristics could become advantageous. At this stage we could expect some organisms to acquire a new type of goal which I shall call "secondary GOALS". these will be learned by experience and transferred to memory. The acquisition of the ability to recognise these - and to steer a pathway towards them, would give any organism a clear survival advantage. The advantage is not merely related to speed of acquisition, but is also depending upon prevailing conditions. For this more advanced method of acquisition of useful characteristics, the research findings of those doing research on REM-dreams are relevant and important (Walker 2017). For example, when sea-living creatures eventually did venture on to that empty land space, it would have been clearly helpful if they had learned what was good to eat, where was a good place to hide and then to build these secondary goals into their regular behaviour. See later for more discussion of this point. A.05 Feature Recognition - using Hard Templates |
|
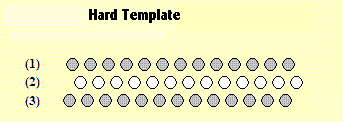
This diagram shows a hard template which can be used to detect a short straight horizontal line segment at one particular location in the visual field. |
|
There are three rows of light sensitive cells in the template which is located within the retina of an eye structure. The top row and the bottom one, are shaded grey. The central row is white. The two grey rows are connected by axons, dendrites and synapse gaps. These connections are not shown in the diagram. The way they are connected, however, ensures that the template operates like this - if darkness impinges on all the cells in that row simultaniously (or on most of them), the whole row (collectively) will emit a positive signal. That is true of both row (1) and row (3). The reverse is true of row (2). If bright light impinges on all (or most of) the cells in row (2), these cells will (collectively) emit a positive signal. These output signals are then analysed and if rows (1 and 3) and row (2) are all emitting positive signals at the same time, then the whole template (collectively) emits a positive signal. Thus the template (as a whole) will emit a positive signal if a bright white horizontal line falls on row (2) while it is also surrounded by darkness. The template is therefore a device which is able to detect a short horizontal white line. At the same time these same cells could be connected in another way so that it will emit a positive signal if and only if a black line segment (with a paler background) impinges upon it. These two forms of connection can exist simultaneusly - side-by-side (as it were). Let us call it a "Hard Template" because it is hard-wired and cannot be changed easily in response to new experience. However, if we want a hard template mechanism to detect short line segments with a range of different locations, different sizes and different orientations, we would need to cover the whole visual field (within the retina) with similar hard templates, in all locations, with a range of sizes and with a range of orientations. That type of arrangement has been found experimentally in the retina of mammals by implanting micro-electrodes into the eyes and the visual cortex of the Cat (Hubel and Wiesel 1957). Similar hard templates can be created for other simple components of visual patterns - edges and corners, for example. A.06 Boundary Classifications |
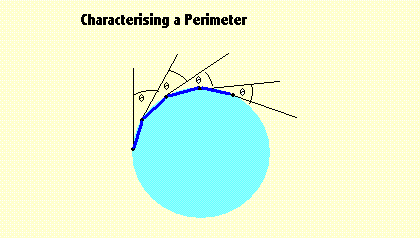
|
This diagram shows a simple circle with component "hard-template" line segments drawn at
regular intervals. Also shown are the differences in angular direction for successive line
segments. If these angles are compared and (within a small tolerance) are found to be equal,
then we can identify the figure as a circle (or a part of a circle). A similar procedure
can be used to identify an open or closed figure, one with a smooth or one with a jagged edge,
a sinusoidal edge, and several more general shapes. The algorithm is inherently slow because
it progresses round the boundary one segment at a time. It cannot start on the next segment
until the previous one has been analysed to yield its location, especially its ending point.
Note: Care must be taken with this algorithm to ensure that a particular figure is not falsely discovered as it might be if the entire region is covered in line segments with different orientations. That possibility must be eliminated by checking that line segments are not present elsewhere. |

| This diagram shows how a partial (or occluded) shape can be recognised using a modification of the same technique. The initial auto-adequacy test would ensure that distinctive partial boundary shapes are present and easily distinguished from the relevant part of the background. If there is no distinctive component or what can be seen cannot easily be distinguished from its background this method of recognition must withdraw itself from the list of alternatives. Note how easily the tail of the cat could have been "lost" among a tangle of branches. In this example however, its shape and position would make a contribution to the overall reliability of the recognition technique. |
|
A.07 Distribution of Area - The Method of Stripes |
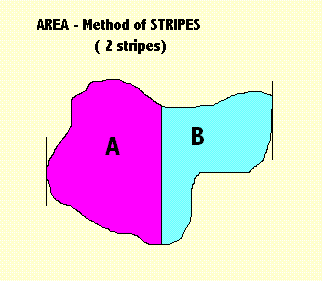
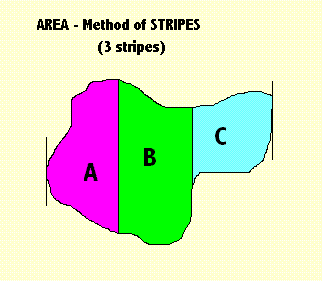
|
|
Before the method can begin, it is necessary to find the perimeter. The method then establishes the two extremities - left and right. 1st Step Divides the area, so bounded, into two parts - "A" and "B". It then calculates the ratio of the areas of these two parts A/B. The diagram above (on the left) illustrates this. 2nd Step Divide the area into three parts. The ratio calculated = B/(A+C). The later steps are obvious: divide the area into 4, 5, 6, 7, .... parts. The choice of ratio does not matter much so long as we are consistent. We compare all shapes using the same ratios. This example also illustrates why, in general, I favour the method of stripes. It is obvious that the method can be extended in easy steps to gradually improve its performance, by being able to discriminate shapes more readily. You can also abridge the algorithm so that (like faces) you can only compare two shapes that are oriented in the same way. This also shows, however, why, as the method improves in performance, it also slows down until it is almost unusable. Conclusion? Find a way to perform these algorithms in parallel. That is, perform them both at the same time. Neither of them has to wait until the other is finished. A.08 Next Stage in the Method of Stripes (Rotation) Next stage of the method rotates the shape by X degrees and repeats that calculation. |
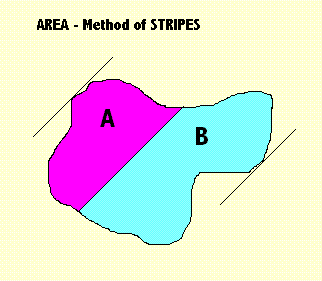
|
In the diagram X = 45 degrees. I have rotated the angle of attack, but in practice the easiest technique is to keep the angle of attack unmoved and to rotate the shape itself by the standard amount. We are off again. The rotation angle X can be made smaller and smaller to get more discrimination between different shapes. More rotations are applied and the same calculation is performed, until a full circle has been completed. A.09 Using Standard Shapes for Comparison |

|
This method is finely tuned by comparing the calculation results performed on an arbitrary shapes (as shown) with the results obtained by applying the same calculations performed on some standard shapes - a circle, a square, an elongated rectangle, a triangle, and so on. We can also introduce some standard natural shapes - a spider, a high-flying bird of prey, a perpetrator and so on. |
|
We can then find - (a) the choice of standard shape and angle of rotation which minimises any discrepancy with one of those standard shapes. (b) the degree of "fit" with the arbitrary standards i.e. is it "roundish", "squarish", "wolfish" and so on? As I indicated this method offers a clear pathway of evolutionary development, from "simple and fast" to "slow and highly accurate" over a the development period. The idea of using pre-calculated characteristics of standard shapes, also appeals because these data can be computed off-line. That saves time (e.g. during dream-sleep). Eventually the stripes can become very narrow and the intervals of rotation can also become very small, to yield a very accurate identification of shape. It occurs to me, that it is the ability to recognise the circumstances in which the methods of recognition fail, that makes the difference between really dependable artificial intelligence and other indifferent methods. |
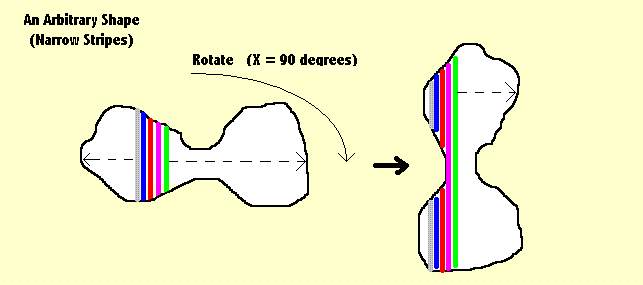
|
The method of stripes operates with arbitrary shapes, but is obviously able to deal best
with silhouettes, for example birds of prey flying high. It does not work well with recognition of
objects against a complicated background.
This diagram offers the example of what we might describe as a "dumbbell" shape. The colours are arbitrary and intended only to discriminate between different stripes. The width of these stripes are variable and can be chosen to suite any given example. They can also be modified as the procedure continues. When the shape divides any given stripe into two (or more) parts we simply add the contribution of each part together. |
|
It can be seen readily enough that what the method is doing, is measuring how the area of the shape is distributed. It does that by determining the numerical ratios between selected pairs of the vertical strips and then ratios of ratios. When the stripes are narrow, a measurement of length is accurate enough to make the measure of area unnecessary. That process can be repeated several times. With each repetition the number of stripes increases, and if necessary, the angle of rotational shift can be decreased. The technique becomes more discriminatory if we also have available a number of standard shapes and the results stored for comparison (see above). We can then compare the results obtained from an arbitrary shape with those obtained by doing the same process on geometrical shapes (a square, a rectangle, a dumbbell, a circle, a star, and so on) we can also include a number of natural shapes (a person, a spider, an eagle, a wolf ...). By comparing the arbitrary shape with each we can find the degree to which it resembles each of the standard shapes and (for each) which orientation provides the best match. A.10 From TWO to THREE Dimensions A logical development from the Method of Stripes takes us to the Method of Columns - as illustrated below. |
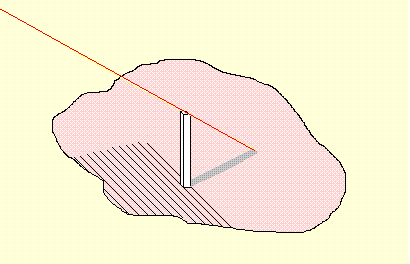
| The diagram shows just a single column. I leave the extention to a picture with many columns to the reader's imagination. The red/orange line is a single ray of light which "explains" the presence of the shadow shown here. |
|
More columns would mean many more rays and many more shadows. Try getting your imagination to build a "column-picture", in the same way, of a human face. I am impressed by the fact that evolution gave up on that and seems not to have tried to give us truly upside-down face templates. This diagram shows only a single narrow ray. Once again we can start with rays which are broad to get an approximate shape and an approximate position of shadows. Rays and columns can also be narrowed progressively to get an increasingly accurate location of shadows. A.11 Feature Recognition and Soft Templates. We saw earlier that it is possible to use what I have called here "Hard Templates" for the recognition of simple elementary components of complicated shapes. The example I used was a short straight line segment. We are able then to have more such hard templates in virtually every location of the visual field, and with more of less all orientations and sizes. What cannot be done, however, is to install a similar range of hard templates which can detect a variety of complex visual patterns (like "faces" for example), with all possible locations, sizes and orientations. Although a typical retina has a very large number of light-sensitive cells, there is simply not a sufficient number of them to accommodate hard templates for the identification of a full range of complex visual patterns. Doing that for all locations, sizes and orientations, demands a supply of resources which is just not available. To obtain an equivalent ability to recognise complicated patterns, we must use an alternative strategy. What we need are "soft templates", which offer, perhaps, only a single exemplar template with just one location, one size and one orientation. The recognition procedure will then need to calculate what are the equivalent of numeric values of location displacement, size magnification/demagnification, and rotational effect required to map the actual signals on to that single fixed template. To store a group of information items like that, one signal is not adequate. What is needed is a cluster of neurones, with interconnections, all of which (collectively) can transmit a cluster of signals. |
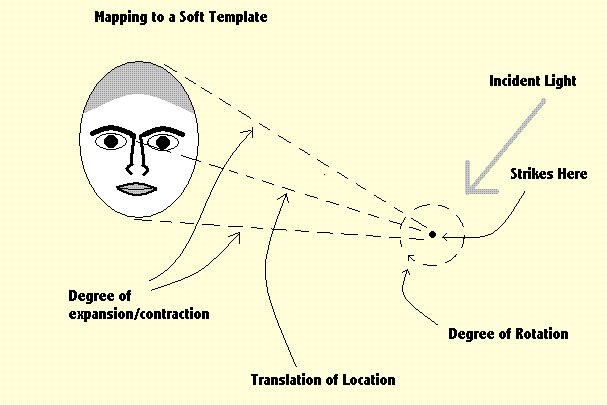
|
The diagram above shows an idealised illustration of an incoming image of a face-like structure, being mapped on to a soft face-template. |
|
In practice several templates would be needed - corresponding to several different views - full frontal, three quarter, profile, etc. With and without hair and/or a beard. The mapping should also reveal the discrepancies discovered, between the standard face-template and actual faces. These discrepancies correspond to relative distance between the eyes etc. (relative to other measures) and it is these discrepancies which can be used to identify individuals. Soft templates must be acquired in childhood with a standard being constructed from the majority of faces observed. If a person grows up among a fairly uniform group of a single racial type, the growing child will not acquire soft templates corresponding to other racial types. This could give rise to the well-known observation that other people of a less frequently observed racial type "all look the same". That is, all people of that type have approximately the same major differences between their faces and the readily available standard template. These mistakes can be corrected later in life, but with increasing difficulty in old age. A.12 Face Recognition Given how important it is for a social animal, to be able to identify a face, first as a face, and then as social contacts become more extensive, being able to identify the individual, the importance of facial recognition is obvious. The use of soft templates for this task is essential but hard to realise. It is perhaps not surprising to find that we have considerable difficulty in recognising the identity of individuals if faces are inverted. It seems clear therefore that we just do not have any templates which are up-side-down even although we can recognise that particular pattern of features is an up-side-down face. Here is an illustration of face recognition and the importance of seeing the relative position of features. 
Now click HERE to see the effect of rearranging those separate (and apparently random collection of dots). If you are reading this text in printout form, turn to Appendix 1 to see the relevant image. A.13 Dealing with Numeric Values To indicate something that is equivalent to a numeric value, I surmise that what is required is a connection to a particular unit in a sequenced order of units. |
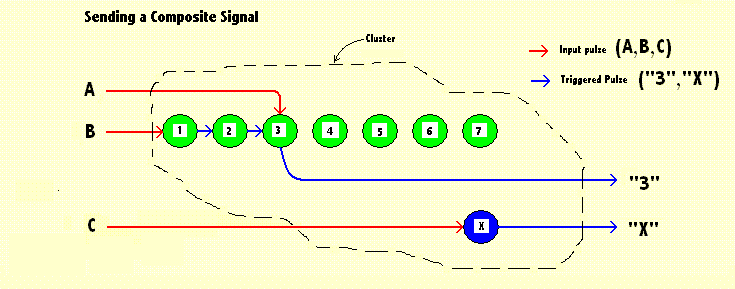
| The diagram above shows a hypothetical arrangement which would enable a cluster of "brain stuff" to send a composite signal to trigger some response. The signal contains an enigmatic "X" and a numerical "3". The arrangement is a conjecture. Other arrangements which could be devised which would enable the same result. This one could operate successfully however. To enable the signals to refer back to their source, these signals would need to be bi-directional (not shown in the diagram). In this example "X" represents whatever designation has been assigned to the source of that signal. It could be many things. The designation would be indicated by the connections to other neurones. |
|
The other part of the output signal shows that the arrangement can also handle numeric data. This can be ordinal numbers and could also be modified slightly to represent cardinal numbers. The array of neurones (coloured GREEN or light GREY) represent the ordinal numbers 1 to 7. Numeric 7 was chosen because psychological tests have shown that that is a representative numeric value which most people can identify at a glance without invoking a counting procedure. That is usually called "subitizing". Input "A" potentiates the 3rd green neurone (or its synaptic connections). Input "B" triggers a procedure which clicks through the array until it reaches the potentiated point where an output signal is then triggered. A.14 Other Perceptual Modalities If we are talking about sound perception, rather than vision, then the RED (or dark grey) sensor blobs on the left of the diagram (in section 2.01) correspond to parts of the cochlea which are sensitive to particular sound frequencies. For other modes of perceptions, however, the round blobs correspond to other types of receptors elsewhere within the living entity. A.15 The Significance of Perceptions The incoming signals are then processed by the associated detection unit. That will be some structure of cells and synapses, equivalent to electronic circuitry representing "AND" and "OR" junctions, which together can detect a particular pattern of signals. Whatever is the type of perception, if the result is positive, an output (or "trigger") signal is sent to an associated action-response unit. I prefer to use the somewhat vague term "unit" because while it will generate (initially) some simple action. But may later evolve into a complex sequence of actions. These will be triggered timeously and in sequence. It may be a muscular action - like running away from danger. It could also be an intellectual procedure - an understanding of the current situation, or merely a rearrangement of stored data. It could also send some pattern of signals to more detection units for further processing. In that way the mechanism could accept a mix of different modes of perception. A.16 Competing Priories What would happen if two separate and quite different events occurred during the same time interval and tried to trigger two different and mutually exclusive action-responses? When that happens the mechanism would need to have a mechanism for dealing with it. The mechanism must be able to decide which of two action-responses takes priority. But this is a simple mechanism which has no intellectual abilities that could be used to choose between alternative actions. There is, however, a way to build an automatic choice into a list of actions that are prepared in advance. Every stimulus-response sub-system must be associated with a priority value, And the one which has the higher priority will be selected (automatically) for action. A.17 Packaged Signals Now ... How can extra information (like a priority value) be added to the messages that are being sent to and fro within a biological brain? This is the problem addressed in the diagram above. That diagram showes how a cluster of so-called "brain stuff" could transmit a composite signal elsewhere within the brain, when it contains numeric data. It identifies a unit within an ordered sequence of units which corresponds to the relevant ordinal number within that sequence. In this case the ordinal sequence corresponds to priority values. The highest priority corresponds to the first in the sequence. In computer science the problem is easily solved, because in a computer the memory store is chopped up into units of a standard size, and because every one of those has a unique numerical "address". we can also form what are called "record structures". These are chunks of computer memory (containing several of these standard units), which can then be stored together contiguously. |
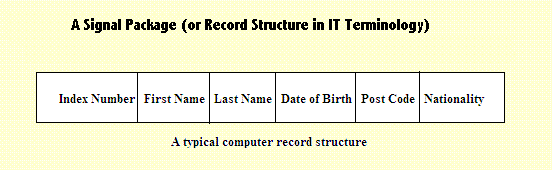
| The diagram above shows a group of data items which refer to a particular person. The data items are the conventional stuff often relating to an individual. |
|
These data can then be used to form, as it were, a package or, if you pefer, an associated bundle, in several adjacent locations. The "adjacent" qualification is important. We can then find all of the data if we know only the main address number (the address number of the first bit of content information). With that we can access all the other contents because they each have a local address (inside the record) rather as individual flats (or apartments) have a small local reference number within a common building. You can, for example, refer to ["Flat No 3 at number 67 Orchard Avenue"]. But how can we associate several separate bits of information with a message which is being transmitted within a biological brain? It is tricky because we do not have a similar access to numerical "addresses". What we do have, however, are a multitude of connections which make use of synapse gaps. These can be switched "ON" and "OFF" by sending pulses to an appropriate location. If all the bits of information which we want to convert into an associated bundle, are already linked by connections which need only the synapse gaps to be switched "ON", then we have a facility which can do the same trick which, as we saw above, can be done within a computer. What is needed then, is an ability to construct a representation of communication between two clusters. But clusters were exactly what had been made available by the construction of concepts - see the description of DOLL_3 below. And that is exactly what I supposed a concept is - a cluster of interconnected neurones and synapses which collectively represent a mental construct of some kind and, collectively, has connections to appropriate and associated other concepts. A cluster of that kind may be a representation of some perceptible physical object, an action-response procedure, or, more generally, just an arbitrary mental construct of some kind which can be used as a structural component when we build a representation of what we suppose is reality. A feature of memory recall, within my own experience, is that these connections to further information, are, as I have said, "made available". But they are not immediately or automatically available. It seems to require the expenditure of energy to render further information, consciously available. That extra effort might correspond to the switching ON of still more synapse connections. That is an expenditure of energy, which some people find unwelcome while for others, doing so may stimulate a degree of pleasure - what psychologists often call the "Ah Ha!" experience. From what has already been noted on that topic, "pleasure" equates to a so-called "scenario" which corresponds to the seeking of a re-experience of something, While the "degree of pleasure" corresponds to the urgency with which it is sought. Ultimately we are all unable to follow these additional connections to very great length. A.18 Recognition of Movement Patterns The ability to detect movement is clearly of great importance. That must also be combined, however, with the ability to distinguish any forms of movement which are important from those which are not important (e.g. caused by the wind), or by a self-induced movement of the body or of the eyes. What we seek, therefore, is a form of perception which can be initially very simple, yet have the potential to develop over a very long time indeed, to become an extremely complicated and subtle mechanism. Of all the possible forms of perception, vision is the only one which has the required properties of ascribing a precise location to individual entities. Other modes of perception are usually restricted the the specification of a direction. The key property of movement is a change of position relative to a fixed environment. When the whole of the environment moves - that can be assigned to the wind or self-movement. At an early stage, therefore, the mechanism of visual perception must evolve an ability to (in effect) "subtract" a general movement of the environment as a whole from an individual perceived movement. There is an added complication. The eye makes small automatic movements called "saccades". There are several explanations for these eye movements - to avoid saturation of any fixed location, to follow extended locations of interest in the visual field, and others. Our interest, however, is concerned with how our evolving mechanism of movement perception could allow for and discount the apparent movement caused by these involontary effects. We need not be overly concerned if our first efforts in the design of these mechanisms, were somewhat crude. They can be improved by pseudo-evolution by gradually adding additional features which will cure individual deficiencies. If we have control over aspects of the relevant environment we could simplify it so that the problems are reduced until our technology is proved adequate. For example, if we want to introduce robot assistants to hospital wards, we could ask all human visitors to wear special vests, or lanyards round their necks (with badges), which can be easily perceived by those robots to ensure that these robots do not collide with them. There should also be control which identify that each visitor is suitably equiped before they enter the wards. These precautions would be abandoned as our robots became more sophisticated. At a very sophisticated level the robots should be able to create an imaginary scene which corresponds to what they ought to be able to perceive. (Now that would represent a really good understanding of events)! When they compare that imaginary scene with what they actually can perceive, they are then able to detect if their perceptions are not adequate. It seems therefore that the ability to imagine a scene is not merely a convenient aid to pleasurable amusement, but provides a genuine and practical survival advantage. It is obvious that techniques for the recognition of characteristic movements, would be a useful additional adjunct for the identification of predators - in forests, in tall grass, and so on. It would also be useful for the recognition of pedestrians, on a road way, in the context of heavy traffic. It is also clear that any creature which is often prey to these predators will usually be able to obtain only a fleeting glimpse of part of that predator's or pedestrian's movements. So how might that be made available? Later, in this text (section 5), I will offer a list of various DOLLS, which would have particular specialisms. One of these will be what I call "DOLL_2" or the "LTSM Memory DOLL". That DOLL, initially, provides the brain mechanism with a specialised memory. "LTSM" stands for "Longer-Term Selective Memory". I have included that to provide a memory of sequences of muscular movements made by the organism itself. I based that proposal on the description given by the animal behaviour expert Konrad Lorenz. He was discussing his observations on shrews in a laboratory observation chamber as they ran about, following a familiar pathway, and also the change of behaviour he saw, when unexpected changes to that pathway were made by himself. The LTSM memory is selective, in that it retains only selected sections of the more primitive TRACE memory. The contents selected for longer retention of which are those which follow a consistent pathway to a condition of particularly high significance. That sequence can then be stored, recalled and re-enacted (or avoided) depending upon the desirability (or otherwise) of that end-condition. Note: the terminology - ".... depending upon the desirability ....". How easily we slip into the terminology of actions being driven by emotional motivation! It is my contention that that expressions of that kind place cause-and-effect the wrong way round. Yet still I fall foul of conventional wording. It occurs to me that the contents of the LTSM memory (or some similar structure) could also provide a memory of short snippets of the patterns of movement seen to be performed by other creatures. Of course, the perception of sounds and scents will also provide a less complicated and perhaps also a more reliable method for the detection of a predator (scent for example), but with the growing importance of vision in a land-based environment, we need to consider how additional visual detection methods could be provided. Once those visible signals were learned and associated with the presence of a predator, they too could be given a high level significance. Evolution has often provided predator animals with a form of camophlage in the form of stripes, which, it seems to me, would be one way of counteracting (by confusion) those remembered snippets of visible patterns of movement. It has also provided those creatures with a style of movement we could describe as "stealthy". A.219 Ships, Shoes and Cabbages Of ships and shoes and sealing wax, and cabbages and kings, And why the sea is boiling hot, and whether pigs have wings. And that brings me to an end of my discussion on methods and procedures for the identification of "things" - i.e. objects, conditions, circumstances et al, that our brain mechanism is likely to encounter in the external (and internal) environment. I am aware that by bringing this particular discussion to an end at this point, I have also left it short, by quite a long way, of the kind of performance achieved by intelligent mammals and, most definitely short of what a human being can achieve. In my own defence, however, I claim that I have set that discussion off to a promising start and that it is travelling in the right direction to achieve those desirable end-points. The mechanisms of a biological brain (i.e. action-potentials travelling along nerve dendrites and axons) are slow acting when compared with electronic pulses travelling along wires (as in a computer). But the brain can compensate for that by being capable of simultaneous or parallel processing. We do have computing machinery that can also do that, but we have not had them for very long. The DOLLS mechanism, which I will begin to discuss from this point onwards, offers a way of examining the potential of that architecture and also a way to segment the problems we will inevitably encounter, and will need to be able to solve. And now I want to consider another question ... what gave me a push towards the idea of the RDM? |